Mouse IL-17/IL-17A Antibody Summary
(rm) IL-17A/IL-17F heterodimer is observed. No cross-reactivity with recombinant human IL-17, recombinant canine IL-17, rmIL-17B,
rmIL‑17C, rmIL-17D, rmIL-17E, or rmIL-17F is observed.
Thr22-Ala158
Accession # Q62386
Applications
Please Note: Optimal dilutions should be determined by each laboratory for each application. General Protocols are available in the Technical Information section on our website.
Scientific Data
 View Larger
View Larger
Detection of Recombinant Mouse IL‑17/IL‑17A by Western Blot. Western blot shows 25 ng of Recombinant Mouse IL-17/ IL-17A (Catalog # 421-ML), Recombinant Human IL-17/IL-17A (Catalog # 317-ILB), Recombinant Rat IL-17/IL-17A (Catalog # 8410-IL), and Recombinant Mouse IL-17F (Catalog # 2057-IL). PVDF Membrane was probed with 1 µg/mL of Rat Anti-Mouse IL-17/ IL-17A Monoclonal Antibody (Catalog # MAB421) followed by HRP-conjugated Anti-Rat IgG Secondary Antibody (Catalog # HAF005). A specific band was detected for IL-17/IL-17A at approximately 15 kDa (as indicated). This experiment was conducted under reducing conditions and using Immunoblot Buffer Group 3.
 View Larger
View Larger
IL‑6 Secretion Induced by IL‑17 and Neutralization by Mouse IL‑17 Antibody. Recombinant Mouse IL-17 (Catalog # 421-ML) stimulates IL-6 secretion in the NIH-3T3 mouse embryonic fibroblast cell line in a dose-dependent manner (orange line), as measured by the Mouse IL-6 Quantikine ELISA Kit (Catalog # M6000B). IL-6 secretion elicited by Recombinant Mouse IL-17 (10 ng/mL) is neutralized (green line) by increasing concentrations of Rat Anti-Mouse IL-17 Monoclonal Antibody (Catalog # MAB421). The ND50 is typically 0.05-0.15 µg/mL.
Reconstitution Calculator
Preparation and Storage
- 12 months from date of receipt, -20 to -70 °C as supplied.
- 1 month, 2 to 8 °C under sterile conditions after reconstitution.
- 6 months, -20 to -70 °C under sterile conditions after reconstitution.
Background: IL-17/IL-17A
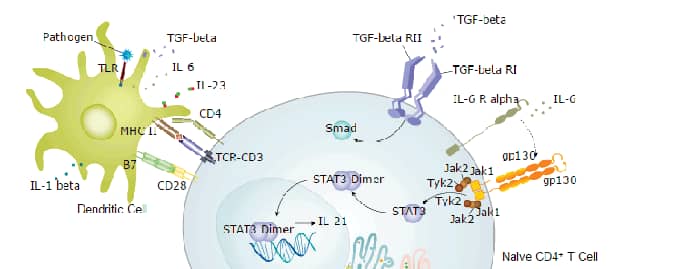
Interleukin 17 (also known as CTLA-8) is a T cell-expressed pleiotropic cytokine that exhibits a high degree of homology to a protein encoded by the ORF13 gene of herpes virus Saimiri. cDNA clones encoding IL-17 have been isolated from activated rat, mouse and human T cells. Mouse IL-17 cDNA encodes a 158 amino acid (aa) residue precursor protein with a 21 amino acid residue signal peptide that is cleaved to yield the 137 aa residue mature IL-17. Both recombinant and natural IL-17 have been shown to exist as disulfide linked homodimers. At the amino acid level, mIL-17 shows 57% and 87% sequence identity with herpesvirus and rat IL-17, respectively. An IL-17 specific mouse cell surface receptor (IL-17 R) has been cloned. While the expression of IL-17 mRNA is restricted to activated alpha beta TCR+CD4-CD8-T cells, the expression of mIL-17 R mRNA has been detected in virtually all cells and tissues tested. IL-17 exhibits multiple biological activities on a variety of cells including: the induction of IL-6 and IL-8 production in fibroblasts; the enhancement of surface expression of ICAM-1 in fibroblasts; activation of NF-kappa B and costimulation of T cell proliferation.
- Kennedy, J. et al. (1996) J. Interferon Cytokine Res. 16:611.
- Yao, Z. et al. (1995) J. Immunol. 155:5483.
- Yao, Z. et al. (1995) Immunity 3:811.
- Rouvier, E. et al. (1993) J. Immunol. 150:5445.
Product Datasheets
Citations for Mouse IL-17/IL-17A Antibody
R&D Systems personnel manually curate a database that contains references using R&D Systems products. The data collected includes not only links to publications in PubMed, but also provides information about sample types, species, and experimental conditions.
91
Citations: Showing 1 - 10
Filter your results:
Filter by:
-
Unraveling CCL20's role by regulating Th17 cell chemotaxis in experimental autoimmune prostatitis
Authors: Zhang, C;Xu, S;Hu, RJ;Liu, XH;Yue, SY;Li, XL;Dai, BS;Liang, CZ;Zhan, CS;
Journal of cellular and molecular medicine
Species: Transgenic Mouse
Sample Types: In Vivo, Whole Cells
Applications: Neutralization -
High glucose intake exacerbates experimental autoimmune prostatitis through mitochondrial reactive oxygen species-dependent TGF-? activation-mediated Th17 differentiation
Authors: Niu, D;Yue, SY;Wang, X;Li, WY;Zhang, L;Du, HX;Liang, CZ;
International immunopharmacology
Species: Murine polyomavirus strain A3
Sample Types: In Vivo
Applications: In vivo assay -
Immunotherapy targeting plasma ASM is protective in a mouse model of Alzheimer's disease
Authors: BJ Choi, MH Park, KH Park, WH Han, HJ Yoon, HY Jung, JY Hong, MR Chowdhury, KY Kim, J Lee, IS Song, M Pang, MK Choi, E Gulbins, M Reichel, J Kornhuber, CW Hong, C Kim, SH Kim, EH Schuchman, HK Jin, JS Bae
Nature Communications, 2023-03-24;14(1):1631.
Species: Mouse
Sample Types: In Vivo
Applications: Neutralization -
Microglial GPR56 is the molecular target of maternal immune activation-induced parvalbumin-positive interneuron deficits
Authors: D Yu, T Li, JC Delpech, B Zhu, P Kishore, T Koshi, R Luo, KJB Pratt, G Popova, TJ Nowakowski, SA Villeda, X Piao
Science Advances, 2022-05-06;8(18):eabm2545.
Species: Mouse
Sample Types: In Vivo
Applications: In Vivo -
Blocking the interaction between interleukin-17A and endoplasmic reticulum stress in macrophage attenuates retinal neovascularization in oxygen-induced retinopathy
Authors: Y Wang, S Gao, S Gao, N Li, B Xie, X Shen
Cell & bioscience, 2021-05-01;11(1):82.
Species: Mouse
Sample Types: Whole Cells
Applications: Neutralization -
T cell-intrinsic role for Nod2 in protection against Th17-mediated uveitis
Authors: RJ Napier, EJ Lee, MP Davey, EE Vance, JM Furtado, PE Snow, KA Samson, SJ Lashley, BR Brown, R Horai, MJ Mattapalli, B Xu, MC Callegan, LS Uebelhoer, CL Lancioni, RK Vehe, BA Binstadt, JR Smith, RR Caspi, HL Rosenzweig
Nat Commun, 2020-10-26;11(1):5406.
Species: Mouse
Sample Types: In Vivo
Applications: Neutralization -
Sex differences in IL-17 contribute to chronicity in male versus female urinary tract infection
Authors: A Zychlinsky, M Rousseau, L Lacerda Ma, T Canton, CR Consiglio, ML Albert, M Fontes, D Duffy, MA Ingersoll
JCI Insight, 2019-05-30;5(0):.
Species: Mouse
Sample Types: In Vivo
Applications: In Vivo -
Endogenous IL-17A mediated neutrophil infiltration by promoting chemokines expression during chlamydial lung infection
Authors: S Qiao, H Zhang, X Zha, W Niu, J Liang, G Pang, Y Tang, T Liu, H Zhao, Y Wang, H Bai
Microb. Pathog., 2019-01-28;0(0):.
Species: Mouse
Sample Types: In Vivo
Applications: Neutralization -
B Cells Produce the Tissue-Protective Protein RELM? during Helminth Infection, which Inhibits IL-17�Expression and Limits Emphysema
Authors: F Chen, W Wu, L Jin, A Millman, M Palma, DW El-Naccach, KE Lothstein, C Dong, KL Edelblum, WC Gause
Cell Rep, 2018-12-04;25(10):2775-2783.e3.
Species: Mouse
Sample Types: In Vivo, Whole Cells
Applications: Flow Cytometry, Neutralization -
Interferon induced protein 35 exacerbates H5N1 influenza disease through the expression of IL-12p40 homodimer
Authors: AP Gounder, CC Yokoyama, NN Jarjour, TL Bricker, BT Edelson, ACM Boon
PLoS Pathog., 2018-04-26;14(4):e1007001.
Species: Mouse
Sample Types: BALF
Applications: Western Blot -
Estrogen Deficiency Promotes Cerebral Aneurysm Rupture by Upregulation of Th17 Cells and Interleukin-17A Which Downregulates E-Cadherin
Authors: BL Hoh, K Rojas, L Lin, HZ Fazal, S Hourani, KW Nowicki, MB Schneider, K Hosaka
J Am Heart Assoc, 2018-04-13;7(8):.
Species: Mouse
Sample Types: In Vivo
Applications: Neutralization -
Kaempferol targeting on the fibroblast growth factor receptor 3-ribosomal S6 kinase 2 signaling axis prevents the development of rheumatoid arthritis
Authors: CJ Lee, SJ Moon, JH Jeong, S Lee, MH Lee, SM Yoo, HS Lee, HC Kang, JY Lee, WS Lee, HJ Lee, EK Kim, JY Jhun, ML Cho, JK Min, YY Cho
Cell Death Dis, 2018-03-14;9(3):401.
Species: Mouse
Sample Types: Cell Culture Supernates
Applications: ELISA Development (Capture) -
Galectin-9 is Critical for Mucosal Adaptive Immunity through the TH17-IgA Axis
Authors: CC Liang, CS Li, IC Weng, HY Chen, HH Lu, CC Huang, FT Liu
Am. J. Pathol., 2018-02-17;0(0):.
Species: Mouse
Sample Types: In Vivo
Applications: Neutralization -
IL-17A Produced by Innate Lymphoid Cells Is Essential for Intestinal Ischemia-Reperfusion Injury
Authors: M Geha, MG Tsokos, RE Bosse, T Sannikova, Y Iwakura, JJ Dalle Lucc, R De Waal Ma, GC Tsokos
J. Immunol., 2017-09-06;0(0):.
Species: Mouse
Sample Types: In Vivo
Applications: Neutralization -
IL-33 Signaling Regulates Innate IL-17A and IL-22 Production via Suppression of Prostaglandin E2 during Lung Fungal Infection
Authors: JM Garth, KM Reeder, MS Godwin, JJ Mackel, CW Dunaway, JP Blackburn, C Steele
J. Immunol., 2017-08-07;199(6):2140-2148.
Species: Mouse
Sample Types: In Vivo
Applications: Neutralization -
Galectin-1-Driven Tolerogenic Programs Aggravate Yersinia enterocolitica Infection by Repressing Antibacterial Immunity
Authors: RC Davicino, SP Méndez-Hue, RJ Eliçabe, JC Stupirski, I Autenrieth, MS Di Genaro, GA Rabinovich
J. Immunol., 2017-07-17;0(0):.
Species: Mouse
Sample Types: In Vivo
Applications: Neutralization -
IL-35 induces N2 phenotype of neutrophils to promote tumor growth
Authors: JM Zou, J Qin, YC Li, Y Wang, D Li, Y Shu, C Luo, SS Wang, G Chi, F Guo, GM Zhang, ZH Feng
Oncotarget, 2017-05-16;8(20):33501-33514.
Species: Mouse
Sample Types: In Vivo
Applications: Neutralization -
Spirulina lipopolysaccharides inhibit tumor growth in a Toll-like receptor 4-dependent manner by altering the cytokine milieu from interleukin-17/interleukin-23 to interferon-?
Authors: H Okuyama, A Tominaga, S Fukuoka, T Taguchi, Y Kusumoto, S Ono
Oncol. Rep, 2017-01-02;37(2):684-694.
Species: Mouse
Sample Types: In Vivo
Applications: Neutralization -
IL-17A exacerbates diabetic retinopathy by impairing M�ller cell function via Act1 signaling
Authors: Ao-Wang Qiu
Exp. Mol. Med, 2016-12-16;48(12):e280.
Species: Mouse
Sample Types: In Vivo
Applications: Neutralization -
IL-17 Augments B Cell Activation in Ocular Surface Autoimmunity
J Immunol, 2016-09-21;0(0):.
Species: Mouse
Sample Types: Whole Cells
Applications: Neutralization -
Interleukin-27 Mediates Susceptibility to Visceral Leishmaniasis by Suppressing the IL-17-Neutrophil Response
Authors: Gustavo F S Quirino
Infect Immun, 2016-07-21;0(0):.
Species: Mouse
Sample Types: In Vivo
Applications: Neutralization -
Seminal plasma induces inflammation in the uterus through the ?? T/IL-17 pathway
Authors: ZH Song, ZY Li, DD Li, WN Fang, HY Liu, DD Yang, CY Meng, Y Yang, JP Peng
Sci Rep, 2016-04-25;6(0):25118.
Species: Mouse
Sample Types: In Vivo
Applications: Neutralization -
IL-17A Is an Important Effector of the Immune Response of the Mammary Gland to Escherichia coli Infection.
Authors: Porcherie A, Gilbert F, Germon P, Cunha P, Trotereau A, Rossignol C, Winter N, Berthon P, Rainard P
J Immunol, 2015-12-18;196(2):803-12.
Species: Mouse
Sample Types: In Vivo, Whole Cells
Applications: Neutralization -
Interleukin-17A and Neutrophils in a Murine Model of Bird-Related Hypersensitivity Pneumonitis.
Authors: Ishizuka M, Miyazaki Y, Masuo M, Suhara K, Tateishi T, Yasui M, Inase N
PLoS ONE, 2015-09-14;10(9):e0137978.
Species: Mouse
Sample Types: In Vivo
Applications: Neutralization -
Dendritic Cells Regulate Treg-Th17 Axis in Obstructive Phase of Bile Duct Injury in Murine Biliary Atresia.
Authors: Liu Y, Li K, Yang L, Tang S, Wang X, Cao G, Li S, Lei H, Zhang X
PLoS ONE, 2015-09-01;10(9):e0136214.
Species: Mouse
Sample Types: In Vivo
Applications: Neutralization -
IL17 Mediates Pelvic Pain in Experimental Autoimmune Prostatitis (EAP).
Authors: Murphy S, Schaeffer A, Done J, Wong L, Bell-Cohn A, Roman K, Cashy J, Ohlhausen M, Thumbikat P
PLoS ONE, 2015-05-01;10(5):e0125623.
Species: Mouse
Sample Types: In Vivo
Applications: Neutralization -
Critical role for IL-18 in spontaneous lung inflammation caused by autophagy deficiency.
Authors: Abdel Fattah E, Bhattacharya A, Herron A, Safdar Z, Eissa N
J Immunol, 2015-04-17;194(11):5407-16.
Species: Mouse
Sample Types: In Vivo
Applications: Neutralization -
Neonatal Streptococcus pneumoniae infection may aggravate adulthood allergic airways disease in association with IL-17A.
Authors: Yang B, Liu R, Yang T, Jiang X, Zhang L, Wang L, Wang Q, Luo Z, Liu E, Fu Z
PLoS ONE, 2015-03-27;10(3):e0123010.
Species: Mouse
Sample Types: In Vivo
Applications: Neutralization -
Regulation of autoimmune germinal center reactions in lupus-prone BXD2 mice by follicular helper T cells.
Authors: Kim Y, Lim H, Jung H, Wetsel R, Chung Y
PLoS ONE, 2015-03-13;10(3):e0120294.
Species: Mouse
Sample Types: Whole Cells
Applications: Neutralization -
Therapeutic efficacy of IL-17A antibody injection in preventing the development of colitis associated carcinogenesis in mice.
Authors: Qi H, Yang H, Xu G, Ren J, Hua W, Shi Y, Torsvik M, Florholmen J, Cui G
Immunobiology, 2014-09-08;220(1):54-9.
Species: Mouse
Sample Types: In Vivo
Applications: Blocking -
Systemic inflammatory response elicited by superantigen destabilizes T regulatory cells, rendering them ineffective during toxic shock syndrome.
Authors: Tilahun A, Chowdhary V, David C, Rajagopalan G
J Immunol, 2014-08-04;193(6):2919-30.
Species: Mouse
Sample Types: In Vivo
Applications: Neutralization -
gammadelta T cells are required for pulmonary IL-17A expression after ozone exposure in mice: role of TNFalpha.
Authors: Mathews J, Williams A, Brand J, Wurmbrand A, Chen L, Ninin F, Si H, Kasahara D, Shore S
PLoS ONE, 2014-05-13;9(5):e97707.
Species: Mouse
Sample Types: In Vivo
Applications: Neutralization -
Porphyromonas gingivalis-derived lysine gingipain enhances osteoclast differentiation induced by tumor necrosis factor-alpha and interleukin-1beta but suppresses that by interleukin-17A: importance of proteolytic degradation of osteoprotegerin by lysine gingipain.
Authors: Akiyama T, Miyamoto Y, Yoshimura K, Yamada A, Takami M, Suzawa T, Hoshino M, Imamura T, Akiyama C, Yasuhara R, Mishima K, Maruyama T, Kohda C, Tanaka K, Potempa J, Yasuda H, Baba K, Kamijo R
J Biol Chem, 2014-04-22;289(22):15621-30.
Species: Bacteria - Porphyromonas gingivalis
Sample Types: Recombinant Protein
Applications: Western Blot -
Leukocyte attraction by CCL20 and its receptor CCR6 in humans and mice with pneumococcal meningitis.
Authors: Klein M, Brouwer M, Angele B, Geldhoff M, Marquez G, Varona R, Hacker G, Schmetzer H, Hacker H, Hammerschmidt S, van der Ende A, Pfister H, van de Beek D, Koedel U
PLoS ONE, 2014-04-03;9(4):e93057.
Species: Mouse
Sample Types: In Vivo
Applications: Neutralization -
Interleukin-17A modulates circulating tumor cells in tumor draining vein of colorectal cancers and affects metastases.
Authors: Tseng J, Yang C, Liang S, Liu R, Yang S, Lin J, Chen Y, Wu Y, Jiang J, Lin C
Clin Cancer Res, 2014-03-27;20(11):2885-97.
Species: Mouse
Sample Types: In Vivo
Applications: Neutralization -
IL-17A produced by neutrophils protects against pneumonic plague through orchestrating IFN-gamma-activated macrophage programming.
Authors: Bi Y, Zhou J, Yang H, Wang X, Zhang X, Wang Q, Wu X, Han Y, Song Y, Tan Y, Du Z, Yang H, Zhou D, Cui Y, Zhou L, Yan Y, Zhang P, Guo Z, Wang X, Liu G, Yang R
J Immunol, 2013-12-13;192(2):704-13.
Species: Mouse
Sample Types: In Vivo
Applications: Neutralization -
CD8 T cells regulate allergic contact dermatitis by modulating CCR2-dependent TNF/iNOS-expressing Ly6C+ CD11b+ monocytic cells.
Authors: Chong S, Tan K, Wong F, Chua Y, Tang Y, Ng L, Angeli V, Kemeny D
J Invest Dermatol, 2013-09-23;134(3):666-76.
Species: Mouse
Sample Types: In Vivo
Applications: Neutralization -
IKKbeta in intestinal epithelial cells regulates allergen-specific IgA and allergic inflammation at distant mucosal sites.
Authors: Bonnegarde-Bernard A, Jee J, Fial M, Aeffner F, Cormet-Boyaka E, Davis I, Lin M, Tome D, Karin M, Sun Y, Boyaka P
Mucosal Immunol, 2013-07-10;7(2):257-67.
Species: Mouse
Sample Types: In Vivo
Applications: Functional Assay -
Galectin-3 negatively regulates dendritic cell production of IL-23/IL-17-axis cytokines in infection by Histoplasma capsulatum.
Authors: Wu S, Yu J, Liu F, Miaw S, Wu-Hsieh B
J Immunol, 2013-03-01;190(7):3427-37.
Species: Mouse
Sample Types: In Vivo
Applications: Neutralization -
Involvement of IL-9 in Th17-associated inflammation and angiogenesis of psoriasis.
Authors: Singh T, Schon M, Wallbrecht K, Gruber-Wackernagel A, Wang X, Wolf P
PLoS ONE, 2013-01-15;8(1):e51752.
Species: Mouse
Sample Types: In Vivo
Applications: Neutralization -
Transcription factors GATA-3 and RORgammat are important for determining the phenotype of allergic airway inflammation in a murine model of asthma.
Authors: Ano S, Morishima Y, Ishii Y, Yoh K, Yageta Y, Ohtsuka S, Matsuyama M, Kawaguchi M, Takahashi S, Hizawa N
J Immunol, 2013-01-04;190(3):1056-65.
Species: Mouse
Sample Types: In Vivo
Applications: Neutralization -
Early IL-17 production by intrahepatic T cells is important for adaptive immune responses in viral hepatitis.
Authors: Hou L, Jie Z, Desai M, Liang Y, Soong L, Wang T, Sun J
J Immunol, 2012-12-10;190(2):621-9.
Species: Mouse
Sample Types: In Vivo, Tissue Homogenates
Applications: Neutralization, Western Blot -
Salmonella enterica induces joint inflammation and expression of interleukin-17 in draining lymph nodes early after onset of enterocolitis in mice.
Authors: Noto Llana M, Sarnacki SH, Vazquez MV, Gartner AS, Giacomodonato MN, Cerquetti MC
Infect. Immun., 2012-04-09;80(6):2231-9.
Species: Mouse
Sample Types: In Vivo
Applications: In Vivo -
IL-23 is required for long-term control of Mycobacterium tuberculosis and B cell follicle formation in the infected lung.
Authors: Khader SA, Guglani L, Rangel-Moreno J
J. Immunol., 2011-10-14;187(10):5402-7.
Species: Mouse
Sample Types: In Vivo
Applications: Neutralization -
TLR2-mediated production of IL-27 and chemokines by respiratory epithelial cells promotes bleomycin-induced pulmonary fibrosis in mice.
Authors: Kim HS, Go H, Akira S, Chung DH
J. Immunol., 2011-09-19;187(8):4007-17.
Species: Mouse
Sample Types: In Vivo
Applications: Neutralization -
Differential effects of peptidoglycan recognition proteins on experimental atopic and contact dermatitis mediated by Treg and Th17 cells.
Authors: Park SY, Gupta D, Kim CH
PLoS ONE, 2011-09-16;6(9):e24961.
Species: Mouse
Sample Types: In Vivo
Applications: Neutralization -
Innate immune responses to systemic Acinetobacter baumannii infection in mice: neutrophils, but not interleukin-17, mediate host resistance.
Authors: Breslow JM, Meissler JJ, Hartzell RR
Infect. Immun., 2011-05-16;79(8):3317-27.
Species: Mouse
Sample Types: In Vivo
Applications: Neutralization -
Contribution of IL-17-producing gamma delta T cells to the efficacy of anticancer chemotherapy.
Authors: Ma Y, Aymeric L, Locher C, Mattarollo SR, Delahaye NF, Pereira P, Boucontet L, Apetoh L, Ghiringhelli F, Casares N, Lasarte JJ, Matsuzaki G, Ikuta K, Ryffel B, Benlagha K, Tesniere A, Ibrahim N, Dechanet-Merville J, Chaput N, Smyth MJ, Kroemer G, Zitvogel L
J. Exp. Med., 2011-03-07;208(3):491-503.
Species: Mouse
Sample Types: Whole Cells
Applications: Neutralization -
A critical function of Th17 proinflammatory cells in the development of atherosclerotic plaque in mice.
Authors: Gao Q, Jiang Y, Ma T, Zhu F, Gao F, Zhang P, Guo C, Wang Q, Wang X, Ma C, Zhang Y, Chen W, Zhang L
J. Immunol., 2010-10-15;185(10):5820-7.
Species: Mouse
Sample Types: In Vivo
Applications: Neutralization -
Molecular immune responses to aerosol challenge with Francisella tularensis in mice inoculated with live vaccine candidates of varying efficacy.
Authors: Shen H, Harris G, Chen W, Sjostedt A, Ryden P, Conlan W
PLoS ONE, 2010-10-12;5(10):e13349.
Species: Mouse
Sample Types: In Vivo
Applications: Neutralization -
Protective role of interleukin-17 in murine NKT cell-driven acute experimental hepatitis.
Authors: Wondimu Z, Santodomingo-Garzon T, Le T, Swain MG
Am. J. Pathol., 2010-09-16;177(5):2334-46.
Species: Mouse
Sample Types: In Vivo
Applications: Neutralization -
Lack of TNFR p55 results in heightened expression of IFN-gamma and IL-17 during the development of reactive arthritis.
Authors: Elicabe RJ, Cargnelutti E, Serer MI, Stege PW, Valdez SR, Toscano MA, Rabinovich GA, Di Genaro MS
J. Immunol., 2010-09-01;185(7):4485-95.
Species: Mouse
Sample Types: In Vivo
Applications: Neutralization -
Th17 immune responses contribute to the pathophysiology of aplastic anemia.
Authors: de Latour RP, Visconte V, Takaku T
Blood, 2010-08-23;116(20):4175-84.
Species: Mouse
Sample Types: In Vivo
Applications: Neutralization -
Enhanced protection to Mycobacterium tuberculosis infection in IL-10-deficient mice is accompanied by early and enhanced Th1 responses in the lung.
Authors: Redford PS, Boonstra A, Read S, Pitt J, Graham C, Stavropoulos E, Bancroft GJ, O'Garra A
Eur. J. Immunol., 2010-08-01;40(8):2200-10.
Species: Mouse
Sample Types: In Vivo
Applications: Neutralization -
Pathological role of interleukin 17 in mice subjected to repeated BCG vaccination after infection with Mycobacterium tuberculosis.
Authors: Cruz A, Fraga AG, Fountain JJ, Rangel-Moreno J, Torrado E, Saraiva M, Pereira DR, Randall TD, Pedrosa J, Cooper AM, Castro AG
J. Exp. Med., 2010-07-12;207(8):1609-16.
Species: Mouse
Sample Types: In Vivo
Applications: Neutralization -
IL-22 defines a novel immune pathway of antifungal resistance.
Authors: De Luca A, Zelante T, D'Angelo C
Mucosal Immunol, 2010-05-05;3(4):361-73.
Species: Mouse
Sample Types: In Vivo
Applications: Neutralization -
IL-17A-producing gammadelta T and Th17 lymphocytes mediate lung inflammation but not fibrosis in experimental silicosis.
Authors: Lo Re S, Dumoutier L, Couillin I, Van Vyve C, Yakoub Y, Uwambayinema F, Marien B, van den Brule S, Van Snick J, Uyttenhove C, Ryffel B, Renauld JC, Lison D, Huaux F
J. Immunol., 2010-04-26;184(11):6367-77.
Species: Mouse
Sample Types: In Vivo
Applications: Neutralization -
Chlamydial respiratory infection during allergen sensitization drives neutrophilic allergic airways disease.
Authors: Horvat JC, Starkey MR, Kim RY, Beagley KW, Preston JA, Gibson PG, Foster PS, Hansbro PM
J. Immunol., 2010-03-12;184(8):4159-69.
Species: Mouse
Sample Types: In Vivo
Applications: Neutralization -
IL-17 produced by neutrophils regulates IFN-gamma-mediated neutrophil migration in mouse kidney ischemia-reperfusion injury.
Authors: Li L, Huang L, Vergis AL, Ye H, Bajwa A, Narayan V, Strieter RM, Rosin DL, Okusa MD
J. Clin. Invest., 2009-12-14;120(1):331-42.
Species: Mouse
Sample Types: In Vivo
Applications: Neutralization -
Encephalitogenicity of complete Freund's adjuvant relative to CpG is linked to induction of Th17 cells.
Authors: Tigno-Aranjuez JT, Jaini R, Tuohy VK, Lehmann PV, Tary-Lehmann M
J. Immunol., 2009-10-07;183(9):5654-61.
Species: Mouse
Sample Types: In Vivo
Applications: Neutralization -
IL-17/Th17 promotes type 1 T cell immunity against pulmonary intracellular bacterial infection through modulating dendritic cell function.
Authors: Bai H, Cheng J, Gao X, Joyee AG, Fan Y, Wang S, Jiao L, Yao Z, Yang X
J. Immunol., 2009-10-07;183(9):5886-95.
Species: Mouse
Sample Types: In Vivo
Applications: Neutralization -
Exaggerated IL-17 response to epicutaneous sensitization mediates airway inflammation in the absence of IL-4 and IL-13.
Authors: He R, Kim HY, Yoon J, Oyoshi MK, MacGinnitie A, Goya S, Freyschmidt EJ, Bryce P, McKenzie AN, Umetsu DT, Oettgen HC, Geha RS
J. Allergy Clin. Immunol., 2009-10-01;124(4):761-70.e1.
Species: Mouse
Sample Types: In Vivo
Applications: Neutralization -
A human colonic commensal promotes colon tumorigenesis via activation of T helper type 17 T cell responses.
Authors: Wu S, Rhee KJ, Albesiano E, Rabizadeh S, Wu X, Yen HR, Huso DL, Brancati FL, Wick E, McAllister F, Housseau F, Pardoll DM, Sears CL
Nat. Med., 2009-08-23;15(9):1016-22.
Species: Mouse
Sample Types: In Vivo
Applications: Neutralization -
Vaccinia virus inoculation in sites of allergic skin inflammation elicits a vigorous cutaneous IL-17 response.
Authors: Oyoshi MK, Elkhal A, Kumar L, Scott JE, Koduru S, He R, Leung DY, Howell MD, Oettgen HC, Murphy GF, Geha RS
Proc. Natl. Acad. Sci. U.S.A., 2009-08-17;106(35):14954-9.
Species: Mouse
Sample Types: In Vivo, Whole Cells
Applications: Neutralization -
Interleukin-17-producing gammadelta+ T cells protect NOD mice from type 1 diabetes through a mechanism involving transforming growth factor-beta.
Authors: Han G, Wang R, Chen G
Immunology, 2009-08-04;129(2):197-206.
Species: Mouse
Sample Types: In Vivo
Applications: Neutralization -
Peroxisome proliferator-activated receptor gamma agonist down-regulates IL-17 expression in a murine model of allergic airway inflammation.
Authors: Park SJ, Lee KS, Kim SR, Min KH, Choe YH, Moon H, Chae HJ, Yoo WH, Lee YC
J. Immunol., 2009-07-29;183(5):3259-67.
Species: Mouse
Sample Types: In Vivo, Tissue Homogenates
Applications: Neutralization, Western Blot -
A MyD88-dependent early IL-17 production protects mice against airway infection with the obligate intracellular pathogen Chlamydia muridarum.
Authors: Zhang X, Gao L, Lei L, Zhong Y, Dube P, Berton MT, Arulanandam B, Zhang J, Zhong G
J. Immunol., 2009-06-19;183(2):1291-300.
Species: Mouse
Sample Types: In Vivo
Applications: Neutralization -
Targeting Tim-1 to overcome resistance to transplantation tolerance mediated by CD8 T17 cells.
Authors: Yuan X, Ansari MJ, D'Addio F, Paez-Cortez J, Schmitt I, Donnarumma M, Boenisch O, Zhao X, Popoola J, Clarkson MR, Yagita H, Akiba H, Freeman GJ, Iacomini J, Turka LA, Glimcher LH, Sayegh MH
Proc. Natl. Acad. Sci. U.S.A., 2009-06-15;106(26):10734-9.
Species: Mouse
Sample Types: In Vivo
Applications: Neutralization -
Tc17, a unique subset of CD8 T cells that can protect against lethal influenza challenge.
Authors: Hamada H, Garcia-Hernandez Mde L, Reome JB, Misra SK, Strutt TM, McKinstry KK, Cooper AM, Swain SL, Dutton RW
J. Immunol., 2009-03-15;182(6):3469-81.
Species: Mouse
Sample Types: In Vivo
Applications: Neutralization -
Interleukin-17 causes neutrophil mediated inflammation in ovalbumin-induced uveitis in DO11.10 mice.
Authors: Zhang Z, Zhong W, Spencer D, Chen H, Lu H, Kawaguchi T, Rosenbaum JT
Cytokine, 2009-02-28;46(1):79-91.
Species: Mouse
Sample Types: In Vivo
Applications: Neutralization -
Anti-inflammatory effects of IL-17A on Helicobacter pylori-induced gastritis.
Authors: Otani K, Watanabe T, Tanigawa T, Okazaki H, Yamagami H, Watanabe K, Tominaga K, Fujiwara Y, Oshitani N, Arakawa T
Biochem. Biophys. Res. Commun., 2009-02-26;382(2):252-8.
Species: Mouse
Sample Types: In Vivo
Applications: Neutralization -
Autoimmunity in dry eye is due to resistance of Th17 to Treg suppression.
Authors: Chauhan SK, El Annan J, Ecoiffier T, Goyal S, Zhang Q, Saban DR, Dana R
J. Immunol., 2009-02-01;182(3):1247-52.
Species: Mouse
Sample Types: In Vivo
Applications: Neutralization -
Promotion of the local differentiation of murine Th17 cells by synovial macrophages during acute inflammatory arthritis.
Authors: Egan PJ, van Nieuwenhuijze A, Campbell IK, Wicks IP
Arthritis Rheum., 2008-12-01;58(12):3720-9.
Species: Mouse
Sample Types: In Vivo
Applications: Neutralization -
Interleukin-1 drives pathogenic Th17 cells during spontaneous arthritis in interleukin-1 receptor antagonist-deficient mice.
Authors: Koenders MI, Devesa I, Marijnissen RJ, Abdollahi-Roodsaz S, Boots AM, Walgreen B, di Padova FE, Nicklin MJ, Joosten LA, van den Berg WB
Arthritis Rheum., 2008-11-01;58(11):3461-70.
Species: Mouse
Sample Types: In Vivo
Applications: Neutralization -
CD8+ Th17 mediate costimulation blockade-resistant allograft rejection in T-bet-deficient mice.
Authors: Burrell BE, Csencsits K, Lu G, Grabauskiene S, Bishop DK
J. Immunol., 2008-09-15;181(6):3906-14.
Species: Mouse
Sample Types: In Vivo
Applications: Neutralization -
Interferon-gamma regulates idiopathic pneumonia syndrome, a Th17+CD4+ T-cell-mediated graft-versus-host disease.
Authors: Mauermann N, Burian J, von Garnier C, Dirnhofer S, Germano D, Schuett C, Tamm M, Bingisser R, Eriksson U, Hunziker L
Am. J. Respir. Crit. Care Med., 2008-05-29;178(4):379-88.
Species: Mouse
Sample Types: In Vivo
Applications: Neutralization -
A distinct subset of natural killer T cells produces IL-17, contributing to airway infiltration of neutrophils but not to airway hyperreactivity.
Authors: Lee KA, Kang MH, Lee YS, Kim YJ, Kim DH, Ko HJ, Kang CY
Cell. Immunol., 2008-04-18;251(1):50-5.
Species: Mouse
Sample Types: In Vivo
Applications: Neutralization -
Lack of Toll IL-1R8 exacerbates Th17 cell responses in fungal infection.
Authors: Bozza S, Zelante T, Moretti S, Bonifazi P, DeLuca A, D'Angelo C, Giovannini G, Garlanda C, Boon L, Bistoni F, Puccetti P, Mantovani A, Romani L
J. Immunol., 2008-03-15;180(6):4022-31.
Species: Mouse
Sample Types: In Vivo
Applications: Neutralization -
Crucial role of the interleukin-6/interleukin-17 cytokine axis in the induction of arthritis by glucose-6-phosphate isomerase.
Authors: Iwanami K, Matsumoto I, Tanaka-Watanabe Y, Inoue A, Mihara M, Ohsugi Y, Mamura M, Goto D, Ito S, Tsutsumi A, Kishimoto T, Sumida T
Arthritis Rheum., 2008-03-01;58(3):754-63.
Species: Mouse
Sample Types: In Vivo
Applications: Neutralization -
Defective tryptophan catabolism underlies inflammation in mouse chronic granulomatous disease.
Authors: D'Angelo C, Segal BH
Nature, 2008-01-10;451(7175):211-5.
Species: Mouse
Sample Types: In Vivo
Applications: Neutralization -
Exacerbation of antigen-induced arthritis in IFN-gamma-deficient mice as a result of unrestricted IL-17 response.
Authors: Irmler IM, Gajda M, Brauer R
J. Immunol., 2007-11-01;179(9):6228-36.
Species: Mouse
Sample Types: In Vivo
Applications: Neutralization -
IL-23 and the Th17 pathway promote inflammation and impair antifungal immune resistance.
Authors: Zelante T, De Luca A, Bonifazi P, Montagnoli C, Bozza S, Moretti S, Belladonna ML, Vacca C, Conte C, Mosci P, Bistoni F, Puccetti P, Kastelein RA, Kopf M, Romani L
Eur. J. Immunol., 2007-10-01;37(10):2695-706.
Species: Mouse
Sample Types: In Vivo
Applications: Neutralization -
Cutting edge: An in vivo requirement for STAT3 signaling in TH17 development and TH17-dependent autoimmunity.
Authors: Harris TJ, Grosso JF, Yen HR, Xin H, Kortylewski M, Albesiano E, Hipkiss EL, Getnet D, Goldberg MV, Maris CH, Housseau F, Yu H, Pardoll DM, Drake CG
J. Immunol., 2007-10-01;179(7):4313-7.
Species: Mouse
Sample Types: In Vivo
Applications: Neutralization -
Epicutaneous antigen exposure induces a Th17 response that drives airway inflammation after inhalation challenge.
Authors: He R, Oyoshi MK, Jin H, Geha RS
Proc. Natl. Acad. Sci. U.S.A., 2007-09-24;104(40):15817-22.
Species: Mouse
Sample Types: In Vivo
Applications: Neutralization -
IL-23 enhances host defense against vaccinia virus infection via a mechanism partly involving IL-17.
Authors: Kohyama S, Ohno S, Isoda A, Moriya O, Belladonna ML, Hayashi H, Iwakura Y, Yoshimoto T, Akatsuka T, Matsui M
J. Immunol., 2007-09-15;179(6):3917-25.
Species: Mouse
Sample Types: In Vivo
Applications: Neutralization -
Resident Vdelta1+ gammadelta T cells control early infiltration of neutrophils after Escherichia coli infection via IL-17 production.
Authors: Shibata K, Yamada H, Hara H, Kishihara K, Yoshikai Y
J. Immunol., 2007-04-01;178(7):4466-72.
Species: Mouse
Sample Types: In Vivo
Applications: Neutralization -
T-bet inhibits both TH2 cell-mediated eosinophil recruitment and TH17 cell-mediated neutrophil recruitment into the airways.
Authors: Fujiwara M, Hirose K, Kagami S, Takatori H, Wakashin H, Tamachi T, Watanabe N, Saito Y, Iwamoto I, Nakajima H
J. Allergy Clin. Immunol., 2007-03-01;119(3):662-70.
Species: Mouse
Sample Types: Whole Cells
Applications: Neutralization -
Functional relevance of the IL-23-IL-17 axis in lungs in vivo.
Authors: Ivanov S, Bozinovski S, Bossios A, Valadi H, Vlahos R, Malmhall C, Sjostrand M, Kolls JK, Anderson GP, Linden A
Am. J. Respir. Cell Mol. Biol., 2006-12-01;36(4):442-51.
Species: Mouse
Sample Types:
Applications: Neutralization -
Interleukin-17 as a recruitment and survival factor for airway macrophages in allergic airway inflammation.
Authors: Sergejeva S, Ivanov S, Lotvall J, Linden A
Am. J. Respir. Cell Mol. Biol., 2005-05-18;33(3):248-53.
Species: Mouse
Sample Types: Whole Cells
Applications: Neutralization -
Inhibition of interleukin-17 prevents the development of arthritis in vaccinated mice challenged with Borrelia burgdorferi.
Authors: Burchill MA, Nardelli DT, England DM, DeCoster DJ, Christopherson JA, Callister SM, Schell RF
Infect. Immun., 2003-06-01;71(6):3437-42.
Species: Mouse
Sample Types: In Vivo
Applications: Neutralization -
Endogenous IL-17 as a mediator of neutrophil recruitment caused by endotoxin exposure in mouse airways.
Authors: Miyamoto M, Prause O, Sjostrand M, Laan M, Lotvall J, Linden A
J. Immunol., 2003-05-01;170(9):4665-72.
Species: Mouse
Sample Types: In Vivo, Whole Cells
Applications: ELISpot Development, ICC, Neutralization
FAQs
-
Does Mouse IL-17/IL-17A antibody (catalog # MAB421) cross-react with Rat IL-17A?
Mouse IL-17/IL-17A antibody, catalog # MAB421, was tested against recombinant Rat IL-17A (catalog # 8410-IL ) in a Direct ELISA assay. 100% cross-rectivity was observed to recombinant Rat IL-17A.
Reviews for Mouse IL-17/IL-17A Antibody
Average Rating: 5 (Based on 2 Reviews)
Have you used Mouse IL-17/IL-17A Antibody?
Submit a review and receive an Amazon gift card.
$25/€18/£15/$25CAN/¥75 Yuan/¥2500 Yen for a review with an image
$10/€7/£6/$10 CAD/¥70 Yuan/¥1110 Yen for a review without an image
Filter by: